Asperoteuthis Nesis, 1980
Asperoteuthis species are deep-living, oceanic squid for which the biology and ecology are poorly known. Recently the first insitu video was made of one squid that produced some surprising information about its biology, especially the function of its elongate secondary fins (see A. mangoldae) which may be typical of all Asperoteuthis spp.
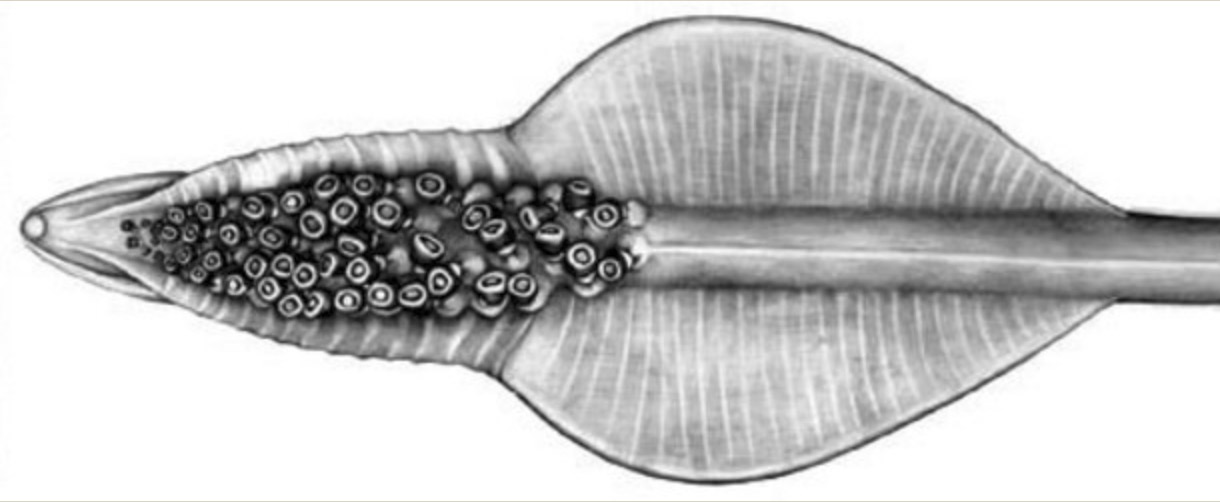
The three known species share tentacles that are somewhat similar to those of Chiroteuthis spp. but are much longer (i.e., A. acanthoderma can have tentacles as much as12 times the mantle length (Tsuchiya and Okutani, 1993)). The clubs of the tentacle that are, also, somewhat similar to those of Chiroteuthis but differ in a way that distinguishes Asperoteuthis spp. from Chiroteuthis spp. as well as all other squid species. The suckers of the Asperoteuthis club exist only in the distal portion of the club while the proximal portion is bare of suckers but has broad protective membranes whose trabeculae are mostly fused.
Unfortunately, due to their long, and slender tentacle stalks the clubs are frequently broken off and lost during capture.
Arms
- Long, generally subequal in length in large subadults with arms IV slightly longer.
- Tentacles
- Club divided into two portions by symmetrical protective membranes.
- Suckers only on distal portion of club.

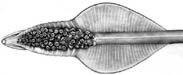
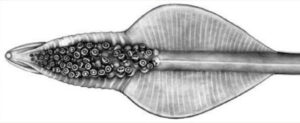
Figure. Oral view of tentacular club of Asperoteuthis acanthoderma. Drawing by J. R. Schroeder. .
- Head
- Olfactory organ located well posterior to each eye.
- Olfactory organ located well posterior to each eye.
- Funnel
- Funnel valve present.
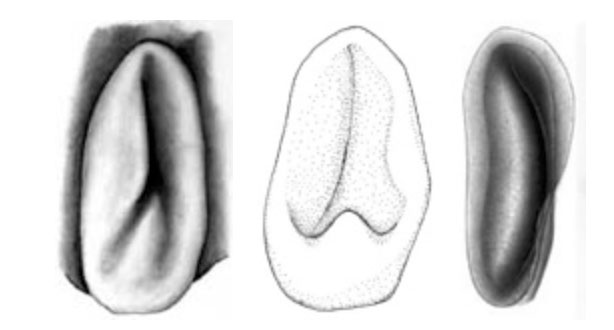
- Funnel component of the funnel-mantle locking-apparatus variable; with inverted Y-shaped groove that poorly defines an elongate tragus and slender antitragus (A. acanthoderma) or a slender antitragus and a strong antitragus (A. lui) or a long tragus (see the light area below) and curved groove (the dark area on the right below) without an antitragus (A. mangoldi).

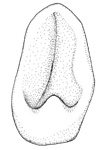
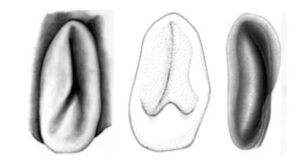
Figure. Ventral view of the funnel locking-apparatus. Left - A. acanthoderma. Drawing by J. R. Schroeder. Middle - A. lui. Drawing from Braid (2016). Right - A. mangoldae. Drawing from Young, et al. (2007). 
- Integument
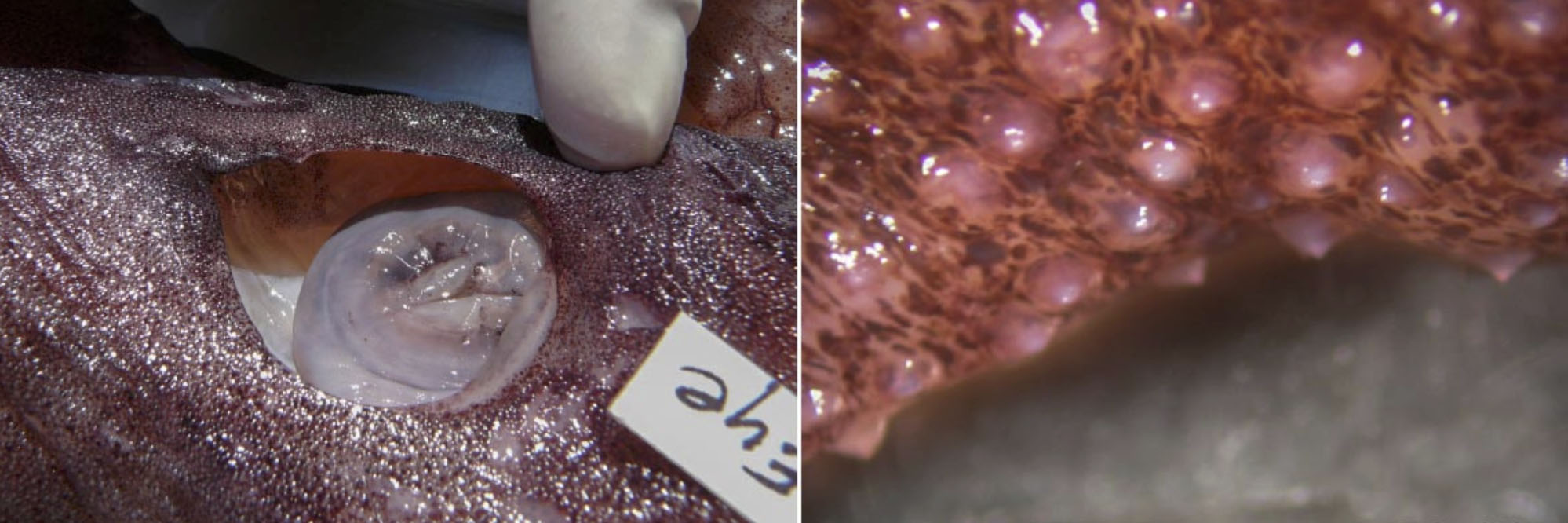
- Skin covered with small tubercules in A. acanthoderma and A. lui but without tubercules in A. mangoldae.
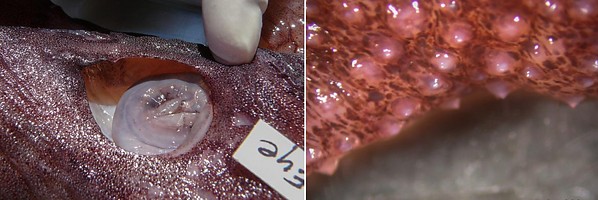
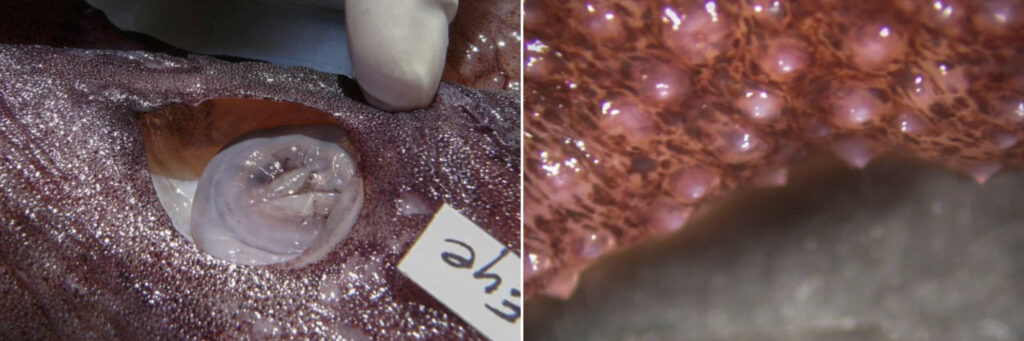
Figure. Integument of A. acanthoderma, mature female, showing tubercules. Photographs by Chuck Stevenson.
- Skin covered with small tubercules in A. acanthoderma and A. lui but without tubercules in A. mangoldae.
- Tail
- Tail with “secondary fin” retained in adults.
- Tail with “secondary fin” retained in adults.
- Photophores
- Photophores absent from viscera and arms IV.
- Large photophore patch on ventral surface of each eyeball.
- Luminescent pads on tentacle stalks (also in Chiroteuthis).
- Aboral surface of club with large, distal photophore (see arrows - terminal position in A. acanthoderma and A. mangoldi, not considering the long, terminal papilla in the latter, well sub-terminal position in A. lui) and a lateral series of small photophores on each side of the aboral club in A. acanthoderma and A. mangoldi (not shown on drawing of latter) and these plus an additional series of midline photophores in A. lui.
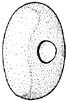
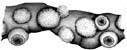
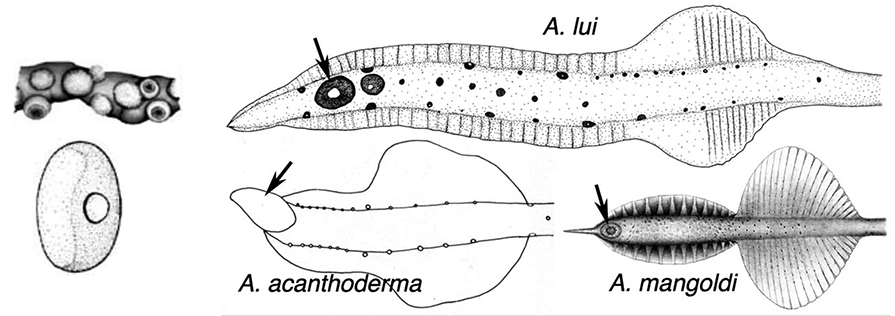
Figures. Left-top - Tentacle stalk photophores, same species. Drawing by J. R. Schroeder. Left-bottom -Ventral view of eye of Asperoteuthis acanthoderma. Drawing from Tsuchiya and Okutani, 1993, showing photophore, with permission. Right - aboral views of tentacle clubs showing photophores. Large distal photophore indicated by arrows. Drawing of A. lui from Braid (2016). Drawing of A. acanthoderma from Lu (1977). Drawing of A. mangoldae from (Young et al., 2007).
Comments
These squid generally lose the tentacles during capture and can easily be confused with Grimalditeuthis (especially A. mangoldae which is of the same size and muscular consistency). However the lack of a fused funnel-mantle locking apparatus easily distinguishes them.
Comparison of major differences between species:
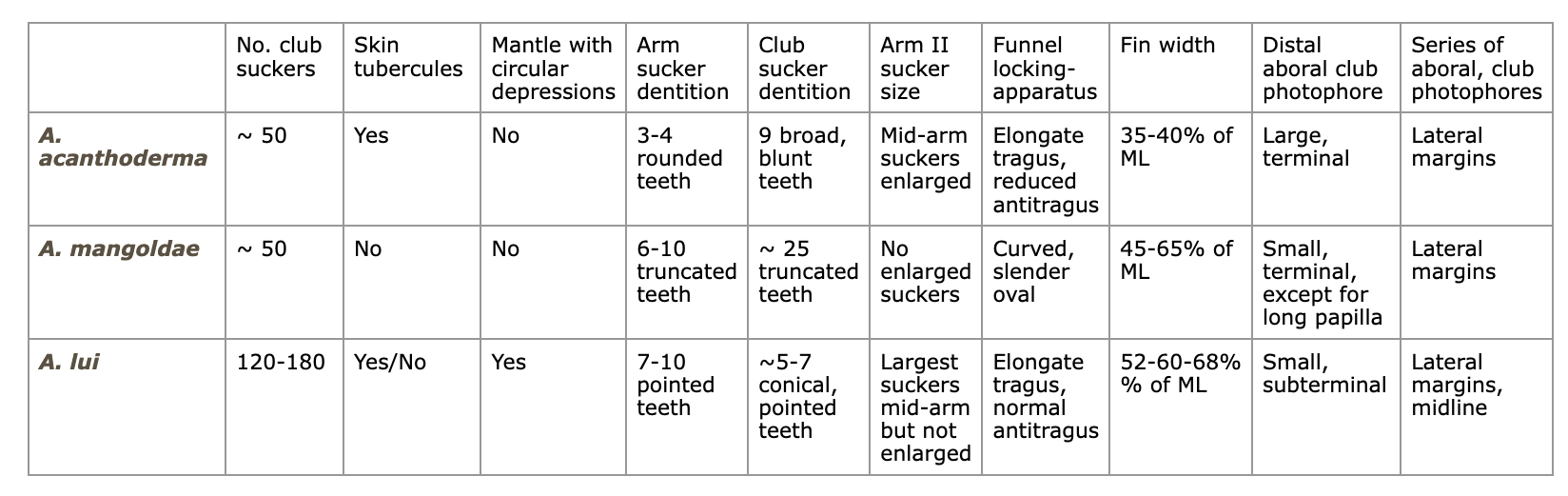
Other characters also separate some or all of the species (e.g., shape of the ocular pphotophore, fin, secondary fin, club and mantle, presence of a long terminal-club papilla).
Paralarvae are known for one species of Asperoteuthis (see Asperoteuthis mangoldae). One figure is added here as it may be critical to the identification of the paralarvae of the other two species:
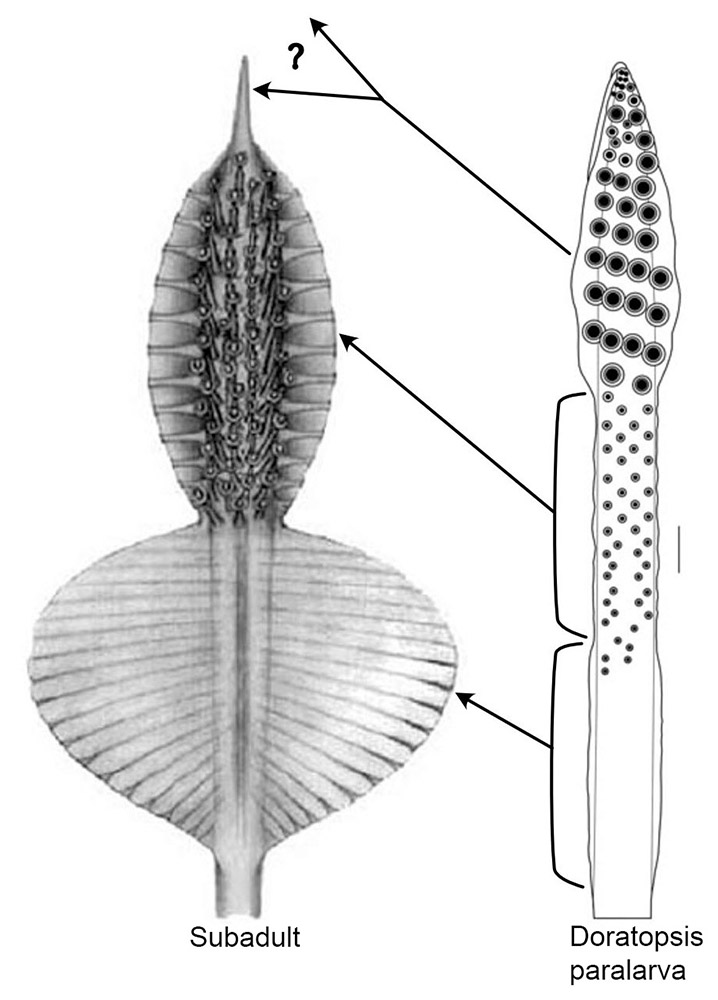
Figure. Left - oral view of the tentacle club of A. mangoldae, subadult, 80 mm ML (Drawing from Young et al. 2007). Right the oral view of the distal part of the tentacle of a A. mangoldae, paralarva, 27 mm ML. (Drawing from Young, 1991).
The arrows indicate the regions of the paralarval tentacle that become the corresponding regions of the subadult club. Top arrow indicates that the distinctive paralarval club becomes either the peculiar long, terminal papilla of the subadult club or is lost completely during metamorphosis. The other two arrows indicate the two regions (which are about the same length) of the developing paralarval tentacle that become most (or all) of the subadult club; upper region is defined by the small, mostly organized, suckers and the low protective membranes; the lower region is defined by the absence of suckers and the distinctively higher protective membranes and their proximal termination.
Lu, C. C. 1977. A new species of squid Chiroteuthis acanthoderma, from the Southwest Pacific (Cephalopoda, Chiroteuthidae). Steenstrupia, 4: 179-188.
Nesis, K. N. 1980. Taxonomic position of Chiroteuthis famelica Berry. Bull. Moscow Obshch. Ispyt. Prirody, sect. Biology, 85: 59-66. [In Russian].
Tsuchiya, K. and T. Okutani. 1993. Rare and interesting squids in Japan -X. Recent occurences of big squids from Okinawa. Venus, 52: 299-311.
Young, R. E. 1978. Vertical distribution and photosensitive vesicles of pelagic cephalopods from Hawaiian waters. Fishery Bulletin, 76: 583-615.
Young, R. E. 1991. Chiroteuthid and related paralarvae from Hawaiian waters. Bull. Mar. Sci., 49: 162-185.
Young, R. E., M. Vecchione and C. F. E. Roper. 2007. A new genus and three new species of decapodiform cephalopods (Mollusca: Cephalopoda). Rev. Fish. Biol. Fisheries, 17: 353-365.